

La palabra inteligente tiene dos significados. Puede aludir simplemente a la actividad de un agente inteligente, aunque el tal actúe de forma estúpida. Por otra parte, puede estar refiriéndose a que un agente inteligente actuó con destreza y maestría. Pasar por alto esta distinción trae como resultado confusión acerca del diseño inteligente. Esta confusión fue llevada hasta mi casa durante una entrevista por radio. El escéptico Michael Shermer y el paleontólogo Donald Prothero me entrevistaron a través de la National Public Radio. Cuando la discusión ya se había desenvuelto, llegue a verme muy sorprendido del significado que ellos le daban a la frase “diseño inteligente”, muy diferente a la acepción dada por la comunidad del diseño inteligente.
Shermer y Prothero malentendieron a la palabra inteligente de “diseño inteligente, dándole un sentido de diseño sabio, astuto o experto. Por consiguiente llegaron a suponer que el diseño inteligente está vinculado a un diseño perfecto. Por otro lado, la comunidad del diseño inteligente entiende al término inteligente de “diseño inteligente” simplemente como referido a una intervención inteligente (independientemente de su destreza o maestría) y por lo tanto separa al diseño inteligente del diseño perfecto.
Entonces ¿cuál es la razón por la que el adjetivo inteligente se encuentra delante del sustantivo diseño? ¿Acaso no está ya incluida en el concepto de diseño la idea de intervención inteligente, convirtiendo a estos dos términos en redundantes? En este caso no hay redundancia debido a que el diseño inteligente necesita ser distinguido, por un lado, del diseño aparente, y por el otro, del diseño perfecto. La frase diseño inteligente refuerza la idea de que el diseño se debe a una inteligencia real, pero también deja totalmente abiertas los atributos o cualidades de esta inteligencia.
En contraste, el diseño aparente plantea que el diseño no es real. Por ejemplo, Richard Dawkins comienza su libro The Blid Watchmaker con la cita, “La biología es el estudio de cosas complejas que dan la apariencia de que han sido diseñadas con un propósito”. Dawkins luego necesita unas trescientas páginas adicionales para argumentar que este diseño es solo una apariencia, y no es real. El diseño aparente, por lo tanto, constituye una negación del diseño inteligente.
Muchos biólogos hacen a un lado al diseño inteligente y a la evidencia a favor de este lanzándose al diseño aparente o al diseño óptimo. Para argumentar del diseño aparente, ellos simplemente apelan al Darwinismo puro y sin ayuda. Para argumentar en contra del diseño inteligente, arrojan argumentos "hombre de paja" con respecto al diseñador, vinculando al diseño inteligente con un diseño perfecto. A fin de dejar al diseño inteligente lo mas implausible que sea posible, ellos definen al diseño perfecto como un diseño óptimo; el mejor, con respecto a todo criterio posible de optimización (presumiblemente, algo de menor calidad no sería digno o esperado de un diseñador inteligente). Como los diseños actuales siempre involucran responsabilidad y compromiso, tales diseños globalmente-óptimos-en-lo-que-a-todo-respecta no puede existir, excepto en un lugar ideal (a veces denominado “Cielo platónico”) muy distante a los diseños reales de este mundo. A diferencia del diseño inteligente, el diseño aparente y el diseño óptimo le quitan al diseño su significado práctico.
Al asimilar que todo diseño biológico es aparente o es óptimo se evita la cuestión central que necesita ser contestada, a saber, si existe actualmente diseño en los sistemas biológicos indiferentemente de los atributos adicionales que estos posean (como el nivel de optimización). Los automóviles que se construyen en plantas de ensamblaje en Detroit están diseñados inteligentemente en el sentido de que una inteligencia humana real es responsable de ello. No obstante, aún si pensáramos que Detroit produce los mejores autos del mundo, nos equivocaríamos al decir que están óptimamente diseñados. Tampoco sería correcto afirmar que están solo aparentemente diseñados (y no porque no estén óptimamente diseñados). ¿Existe alguna razón mínimamente sensata como para que los teóricos del diseño inteligente estén obligados a demostrar que hay diseño óptimo en la naturaleza? Los críticos del diseño inteligente (por ejemplo, Stephen Jay Gould) con frecuencia sugieren que cualquier diseñador cósmico con propósito debería diseñar solo de forma perfecta. Pero esta es una afirmación más teológica que científica.
Aunque atribuir diseño inteligente a artefactos humanos es poco polémico, las cejas se levantan rápidamente cuando el diseño inteligente es atribuido a los sistemas biológicos. Aplicado a la biología, el diseño inteligente sostiene que una inteligencia diseñadora es precisada para diagramar a las estructuras ricas en información y complejas de los organismos vivos. Al mismo tiempo, se niega a especular acerca de la naturaleza de esta inteligencia diseñadora. Mientras que el diseño óptimo demanda la existencia de un diseñador perfeccionista que hace todo siempre de la forma más correcta, el diseño inteligente se adapta a nuestra experiencia ordinaria de diseño, el cual está condicionado por las necesidades de la situación, requiere negociación y responsabilidad, y por consiguiente siempre cae por debajo de un óptimo global e idealizado.

Un diseñador irreal intenta llegar a la perfección en el sentido de lograr el diseño óptimo. En efecto, no existe tal cosa como diseño óptimo. Los diseñadores reales se esfuerzan por lograr una optimización reducida, la cual es algo diferente. Como Henry Petroski, un ingeniero e historiador de la Universidad de Duke, apropiadamente remarca en Invention by Design, “todo diseño implica objetivos conflictivos y por lo tanto compromiso, y los mejores diseños serán siempre aquellos que se inician con el mejor compromiso”. La optimización reducida es el arte del compromiso en medio de objetivos conflictivos. El diseño está basado enteramente en esto. Encontrarle una falla o defecto a un sistema biológico por el hecho de que no cumpla con el estándar óptimo, algo a lo que Gould recurre regularmente, es simplemente facil. Al desconocer los objetivos del diseñador, Gould no se encuentra en una posición de decir si el diseñador propuso un compromiso defectuoso con respecto a los objetivos.
No obstante, la idea de que el diseño inteligente es sub-optimo fue tremendamente exitosa a la hora de detener la discusión acerca del diseño. Curiosamente, este éxito no provino del análisis de alguna estructura biológica determinada, ni de la demostración posterior acerca de cómo la optimización reducida de esta estructura pudo haber sido mejorada. Esto constituiría un análisis científico legítimo, como también que las mejoras propuestas pudieran ser implementadas de forma concreta y no degenerara la ejecución deseada del sistema. Uno puede llegar a imaginar algún perfeccionamiento, pero no tiene idea si podría ser efectuado, o si tal mejora, en el caso de que pueda ser efectuada, no causaría una deficiencia en otra parte.
En mis conferencias públicas pregunté acerca del supuesto diseño sub-optimo del organismo humano. Entre las cosas de las cuales se acusa al diseñador del cuerpo de haber diseñado mal, están la convergencia de la laringe y el esófago en la faringe, que resulta en una gran susceptibilidad a atragantarse; una construcción defectuosa de la espalda que provoca a largo plazo el dolor de espalda, e incluso, en algunos casos de accidentes, la pérdida de motilidad; una pelvis deficiente en las mujeres que trae como consecuencia la dificultad de dar a luz; la pequeñez de la mandíbula humana que resulta en la incapacidad de acomodar a la “dentadura de primate” (la consecuencia más notable se hace ver con los problemas que surgen con la dentadura de juicio). Sin embargo, en el punto más alto de la lista se encuentra la disposición “patas para arriba” del ojo humano. El problema que tienen los biólogos evolucionistas con el ojo humano es que el tal presenta una retina invertida. De acuerdo con esto, los fotorreceptores están orientados en el ojo de tal forma que se encuentran a distancia de la luz entrante y situados por detrás de los nervios y vasos sanguíneos, de los cuales se ha dicho que obstruyen a la luz ingresante.
En realidad, parecen existir buenas razones funcionales para esta construcción. Un sistema visual necesita tres cosas: velocidad, sensibilidad y resolución. La velocidad no se ve afectada por la conexión inversa. También la resolución parece no encontrarse afectada (salvo por una pequeñísima manchita transparente, con la cual el cerebro parece trabajar sin dificultad). En efecto, no existe evidencia de que la retina de los cefalópodos (pulpos y calamares), que se encuentra “correctamente conectada” al tener los receptores orientados hacia adelante y a los nervios plegados hacia atrás, es mejor definiendo objetos en el campo visual. Desde el punto de vista de la sensibilidad, parece que existen buenas razones funcionales para esta inversión de la retina. Las células de la retina son las que más oxigeno necesitan de todas las células del cuerpo humano. Pero ¿en qué momento estas precisan de la mayor cantidad de oxígeno? El requerimiento de oxígeno es el máximo cuando la luz incidente es mínima. El presentar un suministro de sangre en frente de los fotorreceptores garantiza que las células de la retina tengan a su disposición el oxígeno que necesitan para ser tan sensibles como sea posible ante una eventual incidencia mínima de luz. (Algunos vertebrados que presentan a sus retinas invertidas, son sensibles de tal manera que pueden de captar apenas unos pocos fotones).

El diseño es una cuestión de compromiso. No hay ningún problema con que nos guste agregar algo o mejorar a los diseños existentes confiriéndoles funcionalidades adicionales. Sería genial que el ojo humano pudiera gozar de todas sus funcionalidades sin esas minúsculas manchitas imperceptibles. Sería estupendo que el sistema de respiración y el de ingestión conservasen toda su funcionalidad y que a su vez se redujese la incidencia de atragantamiento. Sería muy bueno que nuestras espaldas funcionen como funcionan y que a la vez no nos generen dolor en ciertas situaciónes. Sería lo ideal que la pelvis femenina pudiese funcionar tan bien como funciona y que, sin embargo, se disminuya la dificultad a la hora de parir niños. Sería espléndido que nuestra primera dentadura funcionase perfectamente y no prescindiéramos de dientes y muelas de juicio. Pero cuando se levanta una objeción sobre un diseño acusándolo de ser sub-optimo, invariablemente uno encuentra solo a las funcionalidades adicionales mencionadas, pero no detalles acerca de cómo estas pueden ser implementadas. Y cuando hablamos de diseño, el diablo está en los detalles.
Y aún si los detalles aparecieran, estaríamos caracterizando al diseño solo en base a su calidad (por ejemplo, grado de excelencia). Y aún aquí tenemos que ser cuidadosos. Solo porque el diseño pueda ser mejorado en el sentido de incrementar su funcionalidad en algún aspecto del organismo, eso no significa que tal mejoría será beneficiosa dentro del ancho ecosistema en el cual se encuentra incluido el organismo. La funcionalidad de un depredador puede ser vastamente perfeccionable, pero también puede convertir al predador en uno más dañino para su presa y por consiguiente alterar drásticamente el balance del ecosistema, algo concebiblemente perjudicial para el ecosistema entero. En sus críticas al diseño, los biólogos tienden a colocar su interés sobre las funcionalidades de organismos individuales y ven al diseño como óptimo en el nivel en el cual aquellas funcionalidades se encuentran maximizadas. Pero los diseños elevados de los ecosistemas enteros pueden requerir que los diseños inferiores en los organismos individuales no alcancen su máxima función.
Nuestra visión del diseño también se encuentra muy amoldada por las competencias deportivas. Siempre buscamos ser más rápidos, superiores, altos y fuertes. Pero ¿buscamos realmente ser más rápidos, superiores, altos y fuertes sin un límite? Por supuesto que no. Son precisamente los límites sobre las funcionalidades los que hacen interesante al juego de la vida. (Esta es la razón por la que muchos juegos emplean desventajas). Un jugador de futbol de quinientas libras de peso y con más de 7 piernas, con la fuerza de un gorila y la velocidad de un guepardo sería instantáneamente descalificado de la competencia, porque solo por jugar el juego con la mejor de las habilidades, un jugador tal destrozaría o mataría a los jugadores convencionales que se le crucen por el camino.
Los fans asistirían al juego solo por la novedad o por su deseo de ver sangre, pero un jugador como este destruiría al drama competitivo del deporte. En efecto, este superjugador destruiría o les quitaría a los demás el interés de participar en una competición como esta, y ahí sí que dejaría de ser un juego. Igualmente, un predador como el tal acabaría con todas las presas, y después se extinguiría. O si la supercriatura fuese omnívora y se reprodujera de la forma óptima (como los conejos y las bacterias) hasta terminar con toda la vida, otra vez, después de esto, se extinguiría (a no ser que se convierta en un organismo autótrofo y sintetice su propio alimento, tal como lo hacen algunos organismos unicelulares).
La biología es, entre otras cosas, un drama. Los dramas interesantes requieren de personajes que sean inferiores a los considerados óptimos. De hecho los autores de los dramas humanos diseñan concienzudamente a estos personajes atribuyéndoles defectos y debilidades. ¿Sería Hamlet interesante si Shakespeare no hubiese diseñado al personaje con ciertos defectos y debilidades, tales como su grado de indecisión?
No estoy diciendo que lo defectuoso y lo débil de las características del diseño de los organismos o los ecosistemas sean las bases para realizar una inferencia de diseño. Las inferencias de diseño se diagraman identificando en los sistemas rasgos que sean únicamente diagnostico de diseño. Al mismo tiempo, los desperfectos y las debilidades en las características del diseño de organismos o de ecosistemas pueden ser compatibles con los cambios evolutivos guiados por una inteligencia. En tal escenario evolutivo —en el cual no todos los aspectos de un organismo aislado son considerados óptimos— no se supone que cualquier cambio evolutivo guiado por alguna inteligencia, es necesariamente defectuoso.
Los críticos del diseño inteligente afirman repetidamente que ningún diseñador experto habría creado a todos los callejones sin salida que observamos en el registro fósil. Uno de mis críticos pregunta, ¿qué tiene de inteligente el propósito de crear especies destinadas a la extinción? o ¿por qué un diseñador inteligente crearía humanos con una columna pobremente adaptada para la locomoción bípeda? Si imaginamos a la evolución como progresiva [y esta es una respuesta filosófica propia del autor] en el sentido de que las capacidades de los organismos fueron refinadas y sus humildes orígenes perfeccionados por la selección natural a lo largo del tiempo, entonces parece implausible que un diseñador sabio y benevolente pudiera haber guiado tal proceso. Pero si imaginamos a la evolución como regresiva, en respuesta a una estructura moral distorsionada, resultante de una rebelión humana en contra del diseñador en el punto de inicio temporal, entonces sería posible que un diseñador sin defectos pudiera haberse valido de un proceso evolutivo imperfecto como el medio para inducir al universo pródigo a colocarse bajo su visión de la realidad.
Nos hemos alejado mucho del la cuestión científica, y por una buena razón. Al argumentar que la naturaleza no pudo ser diseñada porque varios sistemas biológicos son sub-óptimos, los oponentes del diseño inteligente han cambiado los términos de la discusión, desde lo científico a lo teológico. En lugar de ¿Cómo puede una estructura existente ser perfeccionada? La pregunta se transforma en, ¿es realmente digno de una deidad crear una estructura como esta? Gould ha sido un experto en el cambio de tema. Por ejemplo, en The Panda´s Thumb, el escribió:
“Si Dios ha diseñado una máquina maravillosa a fin de reflejar su sabiduría y poder, seguramente no se habría valido de una colección de partes usadas generalmente para otros propósitos… las disposiciones desparejas y las soluciones graciosas son la prueba de la evolución —caminos por los cuales un Dios sensible no andaría, pero que los procesos naturales, reducidos por la historia, recorrerían con atrevimiento”.
Aquello que Gould estaba criticando aquí es el “pulgar” del oso panda, una extrusión ósea que le ayuda al panda a desmenuzar el bambú, en particular su dura corteza, y como consecuencia hace que el bambú sea fácil de comer para el panda. (De hecho, el “pulgar” del panda, que en realidad es un sesamoide radial alargado, es extremadamente efectivo a la hora de hacerle fácil la tarea alimenticia al oso).
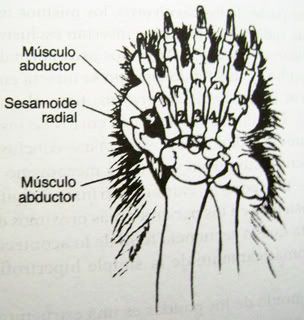
La primera cuestión que necesita ser contestada acerca del pulgar del panda, y en efecto acerca de cualquier estructura biológica, es si esta despliega señales claras de inteligencia. El teórico del diseño no supone que toda estructura biológica es diseñada. Mecanismos naturales como mutación y selección operan en la historia natural para adaptar a los organismos a sus ambientes. Tal vez el pulgar del panda es una adaptación. No obstante, los mecanismos naturales son incapaces de generar estructuras altamente específicas y ricas en información que abundan en la biología. Los organismos despliegan patrones típicos de sistemas tecnológicos inteligentemente pensados —almacenamiento de información y transferencia, códigos funcionales, sistemas de ordenamiento y distribución, autorregulación y ciclos de retroalimentación, circuitos de transducción de señal— y en todas partes, las combinaciones complejas de partes mutuamente interdependientes y bien ensambladas trabajan como en un concierto a fin de desempeñar una función. Los oponentes del diseño inteligente están apegados a equivocaciones, valiéndose de ataques ad hominem, quemando espantapájaros o haciendo afirmaciones teológicas simplistas en lugar de científicas. A lo que no se apegan es a la evidencia pasmante de diseño inteligente, punto que, por lo menos, deberían intentar refutar de una forma lógica.
Autor: William Dembski
-Tiene un Ph.D. en filosofía (Universidad de Illions en Chicago) y un Ph.D. en
matematica (Universidad de Chicago). Es uno de
los principales teóricos del Diseño Inteligente y ha escrito varios libros
sobre la temática. Es autor del primer libro del Diseño Inteligente
publicado por una editorial universitaria renombrada: The Design Inference: Elimitating
Chance Through Small Probabilities. (Cambridge University Press,
1998). Es investigador del Discovery Institute.
Traductor: Daniel Alonso - Estudia
Licenciatura en Ciencias Biológicas en la
UNT , Argentina.








{kind=link}